
Spezies (Arten) von Cannabis
Immer noch herrscht einige Verwirrung über die Zahl der Arten innerhalb der Gattung Cannabis. Einige Botaniker geben aufgrund morphologischer Merkmale drei Arten an: sativa, indica und ruderalis. Sie werden wie folgt charakterisiert: (Beachte, dass Achäne die korrekte Bezeichnung für die Samenform von Cannabis ist. Mit der abgeschrägten Seite sitzt der Same an der Pflanze.)
C. sativa Pflanzen gewöhnlich hoch (150 bis 540 cm), locker verzweigt; Achäne weich; Oberfläche seidig matt, gewöhnlich ohne Marmorierung; fester Sitz am Stiel und ohne bestimmte Ausprägung der Ansaatzstelle.
C. indica: Pflanzen gewöhnlich niedrig (120 cm und weniger); wenig verzweigt, Achäne meist mit marmorierter Oberfläche und ausgeprägter Ansatzstelle; bei Reife leichtes Ausfallen der Samen; Pflanzen sehr dicht verzweigt, mehr oder weniger konisch, gewöhnlich 120 cm hoch und weniger. Ansatzstelle an der Basis der Achäne schwach ausgeprägt.
C. ruderalis: Pflanzen nicht oder kaum verzweigt; gewöhnlich 30-60 cm hoch bei der Reife; Ansatzstelle an der Basis fleischig-runzlig ausgebildet.
C. ruderalis ist eine Wildart, die als Unkraut auf kultivierten Äckern Südost-Rußlands und Zentralasiens auftritt und sich in Richtung Westeuropas ausbreitet. In den USA ist sie nicht zu finden. Auffällig sind die extrem kurzen Blätter und Samen, Samen meist kleiner als die von C. sativa.
C. indica oft mit kleinen, fast runden Samen, die durchschnittlich kleiner als C. ruderalis-Samen bleiben. Pflanze mit wechselständigen Blättern (an jedem Knoten mit 1 Blatt und nach verschiedenen Richtungen zeigend) im Gegensatz zu C. sativa, die meist gegenständige Blätter (2 Blätter stehen sich an jedem Knoten des Stengels gegenüber) aufweist. C. sativa-Samen platzen bei Druckausübung auf den peripheren Grat leicht auseinander, im Gegensatz zu C. indica-Samen. C. sativa bildet die größten Samen der drei Arten aus. Sie sind oft länger als 5 mm. Die Blattspreiten bei
C. sativa sind meist viel schmaler als die von C. indica.
Kreuzungsversuche mit 38 verschiedenen Herkünften zeigten normale Kernteilung, vollständige Austauschbarkeit der Rollen, hohe Pollen-Fertilität und keine Chromosomenmutationen. Dies lässt vermuten, dass keine Fruchtbarkeitsschranken unter den Arten bestehen. C. ruderalis und die wirklich wilden, ursprünglichen Populationen aus Zentralasien (vermutliches Genzentrum) wurden jedoch noch nicht auf ihre Kreuzbarkeit hin untersucht. Ungeachtet der obigen Ergebnisse bilden Fruchtbarkeitsschranken noch kein Kriterium für eine genaue Bestimmung der Arten; die Kriterien liegen eher in einer gleichmäßigen Ausprägung morphologischer und physiologischer Merkmale der Pflanzen, die auch mit hoher Fleritabilität (Vererbbarkeit) an die Nachkommen weitergegeben werden. Um diese Fragen näher klären zu können, sind noch viele Untersuchungen mit den Wildpopulationen aus Zentralasien (wenn sie wirklich existieren) notwendig. Angesichts der verlängerten Keimfähigkeit gefrorener Samen mag es möglich sein, jahrtausende alte Samen in der Arktis zu entdecken, umso den wahren Urahnen des Hanfs einmal zu Gesicht zu bekommen.
Besonders interessant wäre es, ob bestimmte chemische Substanzen (Cannabinoide) in der einen Art auftreten, in den anderen jedoch nicht. Die wenigen Daten, die hierüber erhältlich sind, lassen in dieser Hinsicht nicht viel hoffen. Vier oder fünf als C. ruderalis bestimmte Herkünfte ent-

Abb. 6 Typische Exemplare von Cannabis sativa L.(links), C. indica Lam. (Mitte) und der vermuteten Art C. ruderalis Jan (rechts). Ein typisches Exemplar ist das Exemplar, auf dem die wissenschaftliche Beschreibung einer bestimmten Art basiert. Das „L” nach Cannabis sativa steht für Linn£, der in seinem Werk Systema plantarum (1753) die Namensgebung für Pflanzen zum erstenmal auf eine wissenschaftliche Grundlage stellte. (Nachdruck aus Small, Plant Science Bulletin, vol. 35, 1975)
hielten alle viel CBD und wenig THC, aber weitere Untersuchungen könnten doch lohnenswert sein, da auch bei den anderen beiden Arten Herkünfte mit niedrigen, mittleren und hohen Gehalten der einzelnen Cannabinoide entdeckt wurden. Bis heute ist es noch niemandem gelungen, irgendwelche morphologischen Unterschiede mit chemischen Abweichungen zwischen den Arten zu korrelieren.1
Die Kultivierung von Marihuana mag 10000 Jahre oder mehr zurück- reichen, im Gegensatz zu vielen anderen kultivierten Nutzpflanzen (z.B. Tomaten, Kartoffeln, Mais) wurde es jedoch nie völlig domestiziert. Es entkommt uns leicht und wird wieder zum „Unkraut”. Die Züchtungseffekte in Bezug auf Öl-, Faser- und THC-Gehalt, gepaart mit kontinuierlichem Ausbruch und Kreuzung mit wilden oder domestizierten Populationen haben zu einer extremen Variabilität geführt, die viele spezifische Unterschiede überdecken kann. Schließlich sollte noch bemerkt werden, dass einige Botaniker und auch das Gesetz (in USA) nur die Existenz einer Art anerkennen wollen. Sollte sich diese Ansicht durchsetzen, werden die obigen „Arten” (Species) den Status von Sub species oder Kulturvarietäten einnehmen.
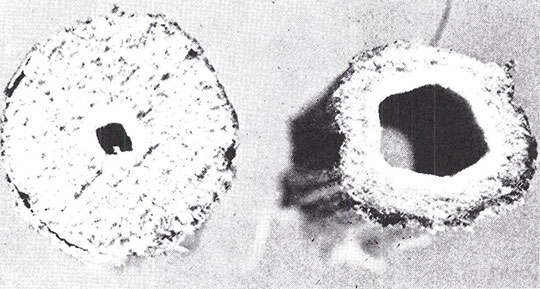
Abb. 7 Querschnitt der Internodien eines Stengels einer THC-armen Faser-Variante (links) und einer THC-reichen Variante (rechts). Faser-Varianten besitzen im Bereich der Internodien einen ausgefüllteren Stengel, der eine höhere Energiezufuhr in das Phloem zwecks höherer Faserproduktion ermöglicht. (Nachdruck aus Small, Plant Science Bulletin, vol. 35, 1975).
Die Samen von Cannabis
Ergänzend zu den oben angeführten Unterschieden zwischen den Samen (korrekter ist die Bezeichnung Früchte) der einzelnen „Arten” sind bleibende Unterschiede zwischen wilden und kultivierten Samen der einzelnen Typen zu nennen. Wilde Samen sind meist kleiner, mit vergleichsweise ausgeprägten Ansatz-Zonen, schmalen Basen und einer papierartigen Außenhaut (Perianth) von marmoriertem oder gesprenkeltem Aussehen. Die Keimung verläuft langsam und unregelmäßig. All diese Charakteristika denkt man sich als Überlebens-Hilfen in der freien Natur.
Wilde Samen werden oftmals nur eine geringe Energiemenge für die Samen-Produktion verwenden, so dass viele kleine Samen offensichtlich einen Selektions- Vorteil bieten. Ebenso helfen die schmalen Basen und gut ausgeprägten Ansatzstellen, die Pflanze weit zu verbreiten, während die marmorierte Oberfläche eine Schutz- oder Abschreck-Funktion gegen Samen-Fresser erfüllt. Bei unregelmäßiger Wasserversorgung wird eine langsame, anpassungsfähige Keimung wünschenswert, besonders in den ersten Monaten der Wachstumsperiode. Viele, wenn nicht nahezu alle, Wild-Populationen sind erst vor kurzem der Kultivierung entflohen. Sie zeigen die obigen Charakteristika in unterschiedlichen Ausprägungen, da sie sich mit ebenfalls verwilderten Drogen- und Faserlinien kreuzten.
Es sollte betont werden, dass es natürlich auch beträchtliche Variationen innerhalb der Linien aus kultivierten und wilden Linien gibt. Eine neuere Studie fand heraus, dass Wildpopulationen im südlichen Asien, besonders in Indien, zum Großteil keinen marmorierten Perianth aufweisen, während die meisten chinesischen wilden Samen ziemlich groß und ohne umhüllenden Perianth oder Basis-Verlängerung ausgeführt sind – sie ähnelten damit Samen kultivierter Pflanzen, die zwar meist den marmorierten Perianth besitzen, welcher aber leicht abfällt und auf reifen Samen selten zu finden ist.
Eine Verallgemeinerung, die man bezüglich Marihuana-Samen treffen kann, ist, dass sie ihre Lebensfähigkeit für extrem lange Zeit behalten, wenn die richtigen Bedingungen herschen. Und die „richtigen” Bedingungen variieren beträchtlich. Ein japanischer Wissenschaftler fand hohe Keimfähigkeit bei Samen, die mit Calciumchlorid 14 Jahre lang trocken gehalten worden waren. Eine andere (amerikanische) Studie über Hanfsamen stellte die tiefen Temperaturen als wichtigsten Faktor heraus: mehr als 95% von Samen, die für 8 Jahre in Stoffbeuteln oder versiegelten Gefäßen gelagert waren, keimten schon bei Temperaturen um 10°C oder darunter. Tiefgefrorene Samen behalten ihre Lebensfähigkeit besonders gut.
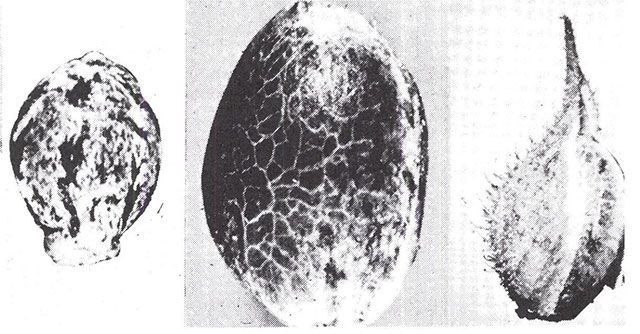
Abb. 8 Samen von Cannabis. Der rechts abgebildete Same ist noch von seinem Perianth umhüllt. (x6). Der linke Same stammt von einer Wildpflanze und der in der Mitte von einer domestizierten Pflanze (beide x9). Beachte, dass der wilde Same kleiner ist, eine schwächer ausgebildete Basis besitzt, seine Ansatzstelle an der Basis gut erkennbar ist und er von einem papierartigen Material, dem Perianth, umgeben ist. (Nachdruck aus Small, Plant Science Bulletin vol. 35, 1975)
Das Trocknen von Samen scheint nur einen geringen Einfluss auszuüben, wenn nicht hohe Temperaturen dabei herrschen. Bei 21°C (70°F) zeigten Samen, die bis zu einem Wassergehalt von 6,2% getrocknet worden waren, keinen Abfall in ihrer Keimfähigkeit nach 5 Jahren, aber Samen, . deren Wassergehalt 9,5% (entspricht etwa dem normalen Feuchtigkeitsgehalt) betrug, zeigten nach 2 Jahren nur noch eine Keimfähigkeit von 5%. Auf alle Fälle behalten Samen, die den normal herrschenden Temperaturen in ungeheizten Gebäuden ausgesetzt sind, nicht ihre optimale Lebensfähigkeit. Aufbewahrt in versiegelten Gefäßen in Kentucky oder Maryland war die Keimungsrate nach 3 Jahren auf 40% abgefallen, im kühlen Klima von Washington war auch nach 6 Jahren noch kein Abfall festzustellen. Es erscheint möglich, dass Samen bei niedrigen oder sehr niedrigen Temperaturen Jahrhunderte überdauern können. Dies würde besonders auf wilde Samen zutreffen, da sie im Durchschnitt sowieso länger lebensfähig bleiben als Samen kultivierter Varietäten. (Alle oben genannten Daten beziehen sich auf Samen des kultivierten Fasertyps von C. sativa).
Zusammenfassung: Von Samen, die in gut versiegelten Gefäßen an einem kühlen Ort wie z.B. im Kühl- oder Gefrierschrank aufbewahrt wurden, kann man eine gute Haltbarkeit erwarten. Wenn sie aber regelmäßig Temperaturen über 50° oder 60°F (auch in verschlossenen Gefäßen) ausgesetzt werden, wird ein markanter Abfall in der Keimfähigkeit schon nach 4 Jahren festzustellen sein, hochgradige Sterilität schon nach 4 Jahren; dies ist zumindest für Samen des CBD-reichen Typs in nördlichen Breiten zutreffend. Solche Keimfähigkeiten können natürlich von Samen, die aus kommerziellem Marihuana stammen, nicht erwartet werden, da solche Samen oft zerquetscht werden oder gar nicht erst die volle Reife erlangen.
Durch selektive Pollen-Befruchtung ist es möglich, Samen zu erhalten, die (fast) nur männliche oder nur weibliche Pflanzen liefern. Dies kann über die Kreuzung bestimmter monözischer (einhäusiger) Linien erreicht werden oder mit der folgenden von fast jedermann durchführbaren Methode: wenn die männlichen Blüten, die aus einer weiblichen Pflanze erwachsen, als Pollenquelle benutzt werden, ergeben die so gewonnenen Samen ausschließlich weibliche Pflanzen. Wenn die Pflanze dies nicht von sich aus tut, muss man verschiedene Techniken anwenden, um diese monözischen Bedingungen zu induzieren. Solche Techniken sind: schwache Beleuchtung, niedrige Nacht-Temperaturen, Stress und Besprühen mit Ethrel. Hierauf wird aber noch in einem späteren Artikel ausführlich eingegangen.
Funktion der Cannabinoide im Marihuana
Es ist ein weit verbreiteter Glaube, dass die Funktion des vom Marihuana ausgeschiedenen Harzes im Schutz vor Austrocknung und im Schutz der Samen besteht. Es ist bekannt, dass Marihuana in nördlichen Breiten oft keinen sichtbaren harzigen Überzug ausbildet, dennoch werden die Samen reif und die Pflanze trocknet nicht aus. In Gegenden, wo die Pflanze freies Harz produziert, kann sie selten durch Trockenheit dazu stimuliert werden; das Harz wird gewöhnlich erst in einem späten Wachstums-Stadium frei, und es sind die jungen Pflanzen, die besonders unter der Tockenheit zu leiden haben. Außerdem scheint keine hohe Korrelation zwischen Harz- Reichtum und hohem THC- oder CBD-Gehalt zu bestehen. Oft findet man Pflanzen mit hohem Cannabinoid-Gehalt, die aber gar nicht besonders harzig oder klebrig sind. Aus diesen Gründen scheinen die Erklärungen für Harz-Produktion und Cannabinoid-Gehalt woanders zu liegen.
Wie bei fast allen anderen psychoaktiven Verbindungen, die in Pflanzen auftreten, ist auch die Funktion der Cannabinoide unbekannt. Gemessen an der Vielzahl der Gene, die für die Synthese solch komplexer Moleküle angelegt werden müssen, darf man wahrscheinlich eine wichtige Bedeutung dieser Moleküle oder ihrer Vorläufer vermuten. Dies macht den Mangel an einer fertigen Erklärung für ihr Auftreten umso frustrierender, besonders, wenn man ein Auge auf die ungeheuren Fortschritte auf dem Gebiet der Pflanzen-Biochemie wirft. Wahrscheinlich bilden die Cannabinoide einen wichtigen Bestandteil des Harzes, welches dazu dienen mag, die Pflanze vor Krankheitserregern, Schadorganismen oder Herbivoren (Pflanzenfressern) zu schützen. Solche sind Bakterien, Viren, Pilze, Insekten und Kühe. Es konnte gezeigt werden, dass Cannabidiol- und Cannabigerol-Säure antibiotisch auf gram-positive Bakterien in vitro (im Reagenzglas) wirken.
Die Gefahr eines pathogenen Angriffs scheint eher in den feucht-heißen Tropen zu bestehen (worauf vielleicht die größere Harzproduktion dort zurückzuführen ist). Dennoch bleibt es ein Geheimnis, warum das THC in südlichen, das CBD in nördlichen Breiten überwiegt.
Seit vielleicht 10000 Jahren haben Menschen Marihuana zur Gewinnung von Fasern, Nahrung (Öl) und Heilmitteln selektiert. Die Varietäten mit intensiver Harz-Produktion wurden sorgfältig gekreuzt und weit verbreitet, und gerade dieses Charakteristikum ist heute wahrscheinlich viel herausragender als in den Hanf-Populationen unserer Vorfahren. Somit mag der Cannabinoid-Reichtum heutiger Marihuana-Pflanzen der Selektion unserer Ahnen zu verdanken sein.
Zellulärer Ursprung der Cannabinoide
Bis zum Erreichen der Blüte besitzen Cannabis-Pflanzen nur zwei Typen von Harzen. Die einzelligen Deckhaare (Trichome) sind lang und dünn und enden in einem Punkt (s. Abb. 9c). Bis zu einem Alter von einem Monat (in warmen Klimaten) oder zwei Monaten (in gemäßigten Klimaten) werden nur wenige dieser Trichome ausgebildet. Später nimmt ihre Zahl gleichmäßig zu, und wenn die Pflanze blüht, bilden sie gewöhnlich an Blüten tragenden Stengeln einen seidigen Flaum. Der zweite Typ von Haaren (Cystolith) ist kurz, geschwollen an der Basis und eingefaßt in die darumliegenden epidermalen Zellen (s. Abb. 9a). Ihr Ende ist stumpf und sie enthalten oft Calciumcarbonat-Kristalle.
Sie werden meist auf der Oberseite von Blättern und Perianthen (kleine spezialisierte Blätter, die die Blüten umgeben) gefunden, manchmal auch auf der Oberfläche von unteren Stengelabschnitten. Wenn die weiblichen Pflanzen kurz vor der Blüte stehen, erhalten ihre Triebspitzen einen Überzug aus vielzelligen Drüsenhaaren, die als kleine schimmernde Punkte sichtbar werden. Bei Beginn der Blüte (?) formen diese Haare eine Basis aus zwei kubischen Zellen, unterstützt durch zwei keilförmige Epidermiszellen und einen globulären Kopf aus vier Zellen, sämtliche überzogen von einer dünnen Kutikula (wachsartige, nicht in Zellen unterteilte Schicht) – (s. Abb. 9f). Zur Zeit der Reife kann der Kopf bis zu 16 radial angeordnete Zellen enthalten, die das Cannabinoid enthaltende Öl als Sekret abscheiden. Es akkumuliert sich dann zwischen den Zellen und der Kutikula (s. Abb. 9b und 9c). Wenn sich genügend Öl angesammelt hat, kann die Kutikula platzen und das Öl (zusammengesetzt aus ungefähr 50% Cannabinoiden) wird frei. Es trocknet dann ziemlich schnell aus und wird zu dem klebrigen Harz. Andere Autoren denken sich die Kutikula selbst aus oxidiertem Harz zusammengesetzt.
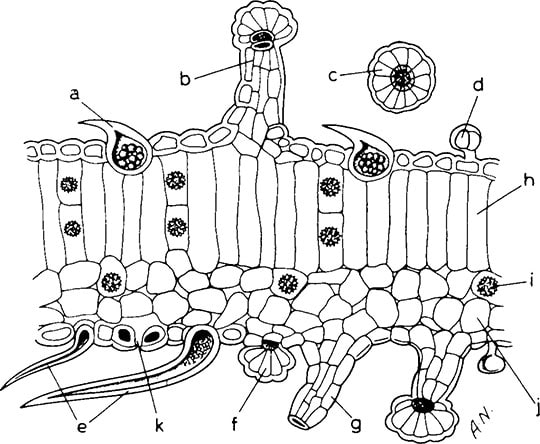
Abb. 9 Querschnitt durch ein Deckblatt einer reifenden Pflanze: a, Cystolith; b, großes Drüsenhaar mit mehreren Zellen in Kopf und Stiel; c, Kopf eines großen Drüsenhaars; d, kleines Drüsenhaar mit zweizeiligem Kopf und einzelligem Stiel; e, dickwandiges konisches Trichom; f, großes, noch unfertiges Drüsenhaar; g, Stiel eines großen Drüsenhaars; h, Zelle des Palisaden-Parenchyms; /, Kristall-Druse;/, Zelle des Schwammparenchyms; k, Spaltöffnung (Nachdruck, mit Erlaubnis, aus Joyce und Curry, The Botany and Chemistry of Cannabis, 1970. Zeichnung von D. Erasmus)
Die beschriebenen Haare variieren in ihrer Erscheinung; Auf Hochblättern und Achsen der weiblichen Blüten sitzende Haare entwickeln einen verlängerten Stiel und ähneln winzigen Pilzen, während andere (besonders auf der Unterseite der oberen Blätter) ungestielt bleiben und einen flachen, runden Kopf aus maximal zehn Zellen (Drüsenschuppen) bilden. Man darf wohl annehmen, dass Drüsenschuppen einen Besonderen Typus und nicht ein frühes Stadium der gestielten Drüsenhaare darstellen. Alle diese Zellen enthalten keine Chloroplasten und sind vom bernsteinfarbigen Öl im Kopf abgesehen farblos. Kleinere Mengen an Cannabinoiden werden auch von Zellen produziert, die im allgemeinen über die gesamte Oberfläche der Epidermis verteilt sind.
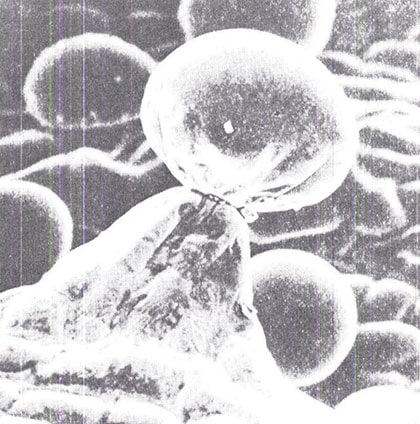
Abb. 10 Elektronenmikioskopische Aufnahme des gestielten Drüsenhaars eines Perianthen in 400facher Vergrößerung. Die Cannabinoide sind im kugeligen Köpfchen konzentriert. Die vorgelagerten „Backsteine” sind Zellen. (Nachdruck aus Fairbairn, Bulletin on Narcotics, vol. 24, 1972).
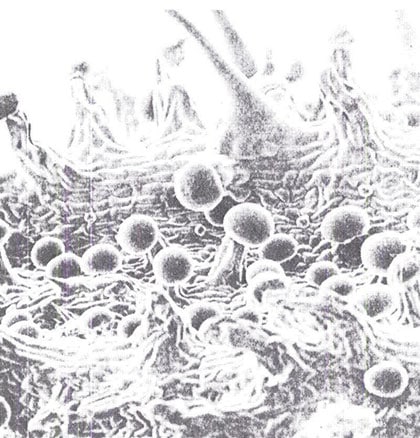
Abb. 11 Elektronenmikroskopische Aufnahme der Blattunterseite eines Perianthen in 100facher Vergrößerung. Die gestielten Drüsenhaare sind in verschiedenen Entwicklungen zu sehen. Oben rechts im Bild sind zwei einfache Deckhaare zu erkennen. (Nachdruck aus Faibairn, Bulletin on Narcotics, vol. 24, 1972)
Vererbung und Umwelt von Cannabis
Ungeachtet der Herkunft eines Samens ähneln die Pflanzen, die an einem anderen Standort aus ihm erwachsen, bereits nach wenigen Generationen den Pflanzen, die an diesem Standort heimisch sind. Diese Tatsache ist seit Jahrhunderten bekannt. Bereits im Jahre 1712 entdeckte Kaempfer, dass Samen aus persischem Cannabis nur noch unbedeutende Mengen berauschenden Harzes produzierten, wenn sie einige Generationen lang in Europa angebaut wurden. Ähnliches entdeckte im frühen 19. Jahrhundert der ägyptische Vizekönig Mehemet Ali, der Hanfsamen aus Frankreich importieren ließ und feststellen musste, dass sie für die Gewinnung von Fasern nicht mehr nütze waren. Nach wenigen Wachstumsperioden wurden die Pflanzen kurz und buschig und produzierten große Mengen an Harz. (Teilweise liegt die Erklärung hierfür auch in der kürzeren Tageslänge in Ägypten). Im gleichen Jahrhundert beobachteten Christison in Edinburgh, Hope in England und Husson in Kairo dasselbe Phänomen an Pflanzen aus importiertem Saatgut. In einem Bericht, der im Jahre 1912 veröffentlicht wurde, beschrieb 1. Bouquet, Pionier der Marihuanaforschung, die Natur dieses Wandels, der in einer einzigen Generation sichtbar wird. Er berichtete, dass Samen aus Indien (Guaza) und Griechenland (Tripolis), die in Frankreich (Lyon) angebaut wurden, Pflanzen von über zwei Metern Höhe ausbildeten. Sie waren robust und buschig, mit vielen Seitentrieben und großen, dunkelgrünen Blättern, die scharf gesägte Ränder und (in der indischen Varietät) deutliche Furchen auf der Oberfläche der Petiolen (Blattstiele) aufwiesen. Die Petiolen der indischen Varietät waren rötlich-braun, verloren aber diese Färbung mit zunehmendem Alter. Die Blüten waren in Büscheln von Trugdolden zusammengefaßt (Haupt- und Nebentriebe enden immer in einer einzelnen Blüte), und die Gruppen der Büschel waren von-einander durch Blätter und gut ausgebildete Brakteen (Deck- oder Trageblätter der Blüten) getrennt. Die kräftigen, großen Blüten gaben einen intensiven Minze-ähnlichen Geruch ab, der noch in zwei Metern Entfernung wahrgenommen werden konnte. Bei der Reife waren die blühenden Spitzen mit leuchtenden Punkten übersät, die aus kleinen rötlichen Harztropfen der Drüsenhaare bestanden.
Als die Samen der obigen Pflanzen im nächsten Jahr in Moulin-sur-Allier ausgesät wurden, ergaben sich Pflanzen, die nur wenig von den in Frankreich zur Fasergewinnung angebauten Hanf-Pflanzen abwichen. Sie waren nur etwas buschiger im Wuchs und eine Spur kräftiger gefärbt als die dort heimischen Pflanzen, besaßen aber nicht mehr die buschigen blühenden Spitzen oder die gefurchten, farbigen Petiolen und die ausgeprägte Harz- Produktion ihrer Vorfahren. Die Spitzen waren nicht einmal klebrig. Im selben Jahr aber wurden die gleichen Samen in Beja, Tunesien (Höhe 330 m ü.N.N.) ausgebracht. Die daraus entstandene Generation war der ersten Generation der indischen und griechischen Pflanzen, die in Lyon gewachsen waren, sehr ähnlich und wies auch die buschigen, duftenden Blüten mit intensiver Harz-Produktion auf.
Die zweite Pflanzen-Generation in Moulin-sur-Allier mag mit den Pflanzen des französischen Faser-Typs Pollen ausgetauscht haben, oder sie war das Ergebnis einer natürlichen Selektion. Es ist wahrscheinlich, dass, wenn die gleichen 100 Samen des indischen Hanfs gleichzeitig in Indien und in Frankreich ausgesät worden wären, Unterschiede in der Keimfähigkeit festzustellen gewesen wären. Ebenso wäre die genetische Ausstattung ihrer Samen sogar nach Inzucht voneinander abweichend gewesen, infolge der unterschiedlichen Selektionskräfte auf sich entwickelnde Samen und Pollen. Die längeren Tage der nördlichen Breiten bewirken wahrscheinlich eine Verlängerung der Internodien und andere Veränderungen im Wuchscharakter der Pflanzen. Später werden noch einige Experimente, die die Effekte unterschiedlicher Tageslängen auf die Harzproduktion prüfen sollen, diskutiert.
Von großem Interesse für das Verständnis der Einwirkungen einer heißen trockenen Umwelt auf die Produktion THC-haltigen Harzes sind die Beobachtungen Bouquets, die er vor etwa 40 Jahren an einer abweichenden Varietät, die 1935 in tunisischen Ernten auftrat, gemacht hat.
Diese Varietät wuchs an zwei voneinander unabhängigen Standorten auf. Die beiden ungefähr 40 km auseinanderliegenden Orte, Jabarka und Sedjenane, sind eingebettet in die Gebirgslandschaft Tunesiens. Wie gewöhnlich war das Saatgut der vorjährigen Ernte entnommen worden, und die Samen, aus denen abnormale Pflanzen entstanden, müssen auf einigen wenigen Pflanzen des Vorjahres gewachsen sein, vielleicht sind sie gar durch Mutation einer einzigen Pflanze entstanden. Besonders auffallend an diesen Pflanzen war ihre äußerst langsame Abreife. Während die normale Varietät binnen 14 Wochen blühte, Samen ansetzte und schon in den ersten Juli-Tagen geerntet werden konnte, hatte der neue Typus auch nach 22 Wochen noch keine Samen angesetzt und wurde Mitte September geerntet. Von größerer Wichtigkeit ist die Tatsache, dass sich diese Merkmale vollständig weitervererbten, und die Pflanzen, die im folgenden Jahr in Tunis und den USA aufwuchsen, im Wesentlichen identisch waren. Es gab weitere auffallende Abweichungen von den normalen Pflanzen. Sie wurden kaum größer als einen Meter, eine Höhe, die für die gewöhnlichen tunesischen Pflanzen durchaus normal ist, aber auch in den amerikanischen Pflanzungen beibehalten wurde, wodurch sie deutlich von den anderen Pflanzen der Population abwichen. Sie waren bis nahe an die Basis reich verzweigt mit einem extrem buschigen, kompakten Habitus. Die Stämme waren allgemein dicker (Stengelumfang bis 9,5 cm an der Basis) und gestauchter als normal, mit längeren, stärkeren Fasern und tieferer Riefung oder Furchung, dennoch fühlten sie sich weicher an; die Pfahlwurzel war rübenförmig (napiform), besaß einen Umfang von etwa 12,2 cm, war 12 cm lang, und gliederte sich weiter auf bis zu einer Gesamtlänge von 32 cm, während der Normal-Typus nicht napiform und allgemein kürzer ausgebildet war. Die neue Varietät war auch mit viel mehr Adventivwurzeln ausgestattet.
Die Blätter waren von der Stammbasis bis zur Spitze durchgehend gegenständig angeordnet, bei der Normal-Varietät manchmal wechselständig. In ihrer Länge (Insertionspunkt an der Petiole bis Ende der mittleren längsten Blattspreite) erreichten sie selten 10 cm, die typischen Blätter dagegen durchschnittlich 20 cm. Alle Blätter waren sieben- oder neunfach gefingert mit 12 bis 16 Zähnen auf jeder Seite, die normalen waren fünf-, sieben-, neun- oder elffach gefingert und zeigten 14 bis 22 Zähne an jeder Seite einer Blattspreite. Die Pflanze weist eine tiefer grüne Färbung der Blattoberseite aber anfänglich hellgrüne Färbung der Blattunterseite auf, die bei der Reife dann fast weißlich erscheint und damit an die Blätter des gemeinen Beifuß (Artemisia vulgaris) erinnert. Die Nervatur der Blätter ist dunkler als normal, so dass man meint, die Nerven wären regelrechte Falten. Sie sind auch von der Blattunterseite betrachtet durchscheinender als gewöhnlich. Die fadenförmigen (filiformen) Nebenblätter (Stipula) an der Basis der Blattstiele (Petiolen) der oberen Blätter sind dunkler gelb-orange gefärbt als normal. Die Brakteen (spezialisierte Blättchen, die die Blüte um geben) sind besser ausgebildet, länger und zahlreicher als gewöhnlich und bilden kleine Büschel. Die blühenden Spitzen sind weniger zahlreich, länger oder weniger kompakt als typisch mit 0 bis 8 Samen pro Infloreszenz, verglichen mit der normalen Anzahl von 40 bis 60. Die Spitzen sind weder klebrig, noch weisen sie den Minze-Geruch auf, selbst wenn sie zwischen den Fingern zerrieben werden.
Bei der mikroskopischen Betrachtung erscheint das Puder von den Rändern der Blattspreiten eher bräunlich-grün als gelb-grün, und wiederum fehlt der charakteristische Geruch des frischen Puders. Die Mikrostruktur ist normal, wir sehen doppelte Calciumoxalat-Kristalle und verschiedene Typen von Haaren: einzellig-punktförmige, kurze Cystolithen (mit Calciumcarbonat-Kristallen im Innern), konisch geschwollen und mit stumpfen Ausläufern etc., die keulenartigen, gestielten (pedizellulären) sekretorischen Haare sind jedoch selten, zudem wenig entwickelt und enthalten nicht das klare bernsteinfarbene Harz. Männliche und weibliche blühende Spitzen zeigen positive Reaktionen für Cannabidiol. Die ..männliche” Reaktion fällt aber schwächer aus und gleicht der „weiblichen” Reaktion vor der Blüte.
Diese Beobachtungen zeigen, dass wir nicht zwingend erwarten können, dass Pflanzen in ihrer Erscheinung oder ihrem THC-Gehalt genau wie ihre Eltern ausfallen, nicht einmal in der ersten Generation. In den nachfolgenden Generationen können die Unterschiede noch größer werden, bis die Pflanzen schließlich kaum mehr von den heimischen unterscheidbar sind. Wenn wir aber besonders umsichtig bei der Wahl unserer Pollenquelle sind, können wir vielleicht mehrere Generationen lang THC-reiche Linien erzeugen, denn wie die nächsten Artikeln zeigen werden, ist die Intensität der Cannabinoid-Produktion in erster Linie genetisch fixiert.
Bonsai-Marihuana
Im allgemeinen gehört Marihuana zu den einjährigen Gewächsen. Es blüht und stirbt jedes Jahr aufs Neue. In der freien Natur trifft dies wahrscheinlich immer zu, auch in warmen Klimaten. (Natürlich können in ganzjährig warmen Gebieten ständig neue Pflanzen entstehen. Es existieren Berichte, nach denen C. sativa in manchen tropischen und subtropischen Gegenden perenniert (mehrjährig wird). Nichtsdestoweniger kann man wahrscheinlich durch geeignete Manipulation Pflanzen heranziehen, die Jahre oder Jahrzehnte alt werden. Einige Leute haben weibliche Pflanzen im Freien mehrere Jahre lang kultiviert, indem sie ständig die blühenden Spitzen abkürzten, aber langlebige Pflanzen können wohl eher im Haus gehalten werden, wo die Umweltbedingungen besser zu kontrollieren sind. Jemand in Los Angeles behauptet, er habe eine Marihuana-Pflanze in seinem Haus, die seit mehr als 3 Jahren ein überaus potentes Produkt abwirft. Inwieweit die traditionellen Bonsai-Techniken wie Beschneiden, Beschränkung von Wasser- und Nährstoffgaben etc., auf Marihuana anwendbar sind, ist noch unbekannt. Auf diesem Gebiet sind noch sorgfältige Versuche notwendig.





